Q9FF14
Gene name |
FH19 (At5g07780, MXM12.2) |
Protein name |
Formin-like protein 19 |
Names |
AtFH19 |
Species |
Arabidopsis thaliana (Mouse-ear cress) |
KEGG Pathway |
ath:AT5G07780 |
EC number |
|
Protein Class |
|
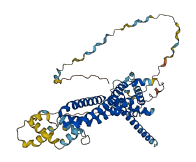
Descriptions
The autoinhibited protein was predicted that may have potential autoinhibitory elements via cis-regPred.
Autoinhibitory domains (AIDs)
Target domain |
|
Relief mechanism |
|
Assay |
cis-regPred |
Accessory elements
No accessory elements
Autoinhibited structure

Activated structure
1 structures for Q9FF14
| Entry ID | Method | Resolution | Chain | Position | Source |
|---|---|---|---|---|---|
| AF-Q9FF14-F1 | Predicted | AlphaFoldDB |
53 variants for Q9FF14
| Variant ID(s) | Position | Change | Description | Diseaes Association | Provenance |
|---|---|---|---|---|---|
| tmp_5_2479714_T_C | 3 | L>P | No | 1000Genomes | |
| ENSVATH03035766 | 10 | Y>D | No | 1000Genomes | |
| ENSVATH03035766 | 10 | Y>N | No | 1000Genomes | |
| tmp_5_2479744_T_C | 13 | V>A | No | 1000Genomes | |
| tmp_5_2479860_C_T | 52 | R>C | No | 1000Genomes | |
| tmp_5_2479891_C_T | 62 | P>L | No | 1000Genomes | |
| ENSVATH10642449 | 64 | L>R | No | 1000Genomes | |
| ENSVATH06938570 | 67 | P>L | No | 1000Genomes | |
| ENSVATH06938571 | 74 | T>A | No | 1000Genomes | |
| tmp_5_2479956_G_C | 84 | V>L | No | 1000Genomes | |
| ENSVATH10642451 | 93 | S>T | No | 1000Genomes | |
| ENSVATH00611345 | 103 | E>A | No | 1000Genomes | |
| ENSVATH03035771 | 121 | S>P | No | 1000Genomes | |
| tmp_5_2480195_G_A | 124 | E>K | No | 1000Genomes | |
| ENSVATH03035772 | 126 | I>L | No | 1000Genomes | |
| ENSVATH03035773 | 129 | L>V | No | 1000Genomes | |
| ENSVATH03035775 | 134 | K>R | No | 1000Genomes | |
| ENSVATH03035776 | 138 | E>K | No | 1000Genomes | |
| ENSVATH00611346 | 140 | E>Q | No | 1000Genomes | |
| tmp_5_2480652_A_C | 190 | N>T | No | 1000Genomes | |
| ENSVATH03035781 | 195 | K>E | No | 1000Genomes | |
| ENSVATH13956029 | 196 | E>K | No | 1000Genomes | |
| tmp_5_2480968_A_G | 216 | Q>R | No | 1000Genomes | |
| ENSVATH03035784 | 243 | Q>K | No | 1000Genomes | |
| tmp_5_2481190_A_G | 245 | T>A | No | 1000Genomes | |
| tmp_5_2481193_A_G | 246 | K>E | No | 1000Genomes | |
| tmp_5_2481200_C_A | 248 | T>N | No | 1000Genomes | |
| tmp_5_2481227_C_G | 257 | A>G | No | 1000Genomes | |
| ENSVATH03035789 | 286 | G>A | No | 1000Genomes | |
| ENSVATH06938582 | 289 | R>S | No | 1000Genomes | |
| ENSVATH10642569 | 326 | S>A | No | 1000Genomes | |
| tmp_5_2481813_A_T | 333 | K>* | No | 1000Genomes | |
| ENSVATH03035804 | 336 | E>Q | No | 1000Genomes | |
| ENSVATH06938583 | 337 | S>N | No | 1000Genomes | |
| ENSVATH03035812 | 345 | Q>H | No | 1000Genomes | |
| ENSVATH03035811 | 345 | Q>K | No | 1000Genomes | |
| ENSVATH10642595 | 346 | L>V | No | 1000Genomes | |
| ENSVATH06938585 | 354 | Q>H | No | 1000Genomes | |
| ENSVATH03035814 | 355 | A>D | No | 1000Genomes | |
| ENSVATH06938586 | 356 | I>V | No | 1000Genomes | |
| tmp_5_2481979_G_T | 364 | K>N | No | 1000Genomes | |
| ENSVATH03035815 | 367 | L>I | No | 1000Genomes | |
| ENSVATH03035817 | 372 | T>S | No | 1000Genomes | |
| tmp_5_2482235_G_A | 399 | M>I | No | 1000Genomes | |
| tmp_5_2482520_C_G | 426 | T>S | No | 1000Genomes | |
| ENSVATH10642599 | 427 | A>T | No | 1000Genomes | |
| tmp_5_2482531_T_A | 430 | L>M | No | 1000Genomes | |
| tmp_5_2482559_C_G | 439 | A>G | No | 1000Genomes | |
| tmp_5_2482579_C_G | 446 | Q>E | No | 1000Genomes | |
| ENSVATH03035819 | 448 | E>D | No | 1000Genomes | |
| tmp_5_2482603_G_A | 454 | A>T | No | 1000Genomes | |
| tmp_5_2482613_A_T | 457 | E>V | No | 1000Genomes | |
| ENSVATH06938593 | 460 | M>L | No | 1000Genomes |
No associated diseases with Q9FF14
1 regional properties for Q9FF14
| Type | Name | Position | InterPro Accession |
|---|---|---|---|
| domain | Formin, FH2 domain | 68 - 464 | IPR015425 |
No GO annotations of cellular component
| Name | Definition |
|---|---|
| No GO annotations for cellular component |
1 GO annotations of molecular function
| Name | Definition |
|---|---|
| actin binding | Binding to monomeric or multimeric forms of actin, including actin filaments. |
2 GO annotations of biological process
| Name | Definition |
|---|---|
| actin nucleation | The initial step in the formation of an actin filament, in which actin monomers combine to form a new filament. Nucleation is slow relative to the subsequent addition of more monomers to extend the filament. |
| barbed-end actin filament capping | The binding of a protein or protein complex to the barbed (or plus) end of an actin filament, thus preventing the addition, exchange or removal of further actin subunits. |
12 homologous proteins in AiPD
| UniProt AC | Gene Name | Protein Name | Species | Evidence Code |
|---|---|---|---|---|
| A0A1D5P556 | DAAM2 | Disheveled-associated activator of morphogenesis 2 | Gallus gallus (Chicken) | SS |
| O95466 | FMNL1 | Formin-like protein 1 | Homo sapiens (Human) | SS |
| Q27J81 | INF2 | Inverted formin-2 | Homo sapiens (Human) | EV |
| Q9Y4D1 | DAAM1 | Disheveled-associated activator of morphogenesis 1 | Homo sapiens (Human) | EV |
| Q86T65 | DAAM2 | Disheveled-associated activator of morphogenesis 2 | Homo sapiens (Human) | SS |
| Q80U19 | Daam2 | Disheveled-associated activator of morphogenesis 2 | Mus musculus (Mouse) | SS |
| Q8BPM0 | Daam1 | Disheveled-associated activator of morphogenesis 1 | Mus musculus (Mouse) | PR |
| Q0GNC1 | Inf2 | Inverted formin-2 | Mus musculus (Mouse) | SS |
| Q0QWG9 | Grid2ip | Delphilin | Mus musculus (Mouse) | PR |
| Q9C7S1 | FH12 | Formin-like protein 12 | Arabidopsis thaliana (Mouse-ear cress) | PR |
| Q9LH02 | FH17 | Formin-like protein 17 | Arabidopsis thaliana (Mouse-ear cress) | PR |
| P0C5K4 | FH21A | Putative formin-like protein 21a | Arabidopsis thaliana (Mouse-ear cress) | PR |
| 10 | 20 | 30 | 40 | 50 | 60 |
| MSLVDISGAY | SLVPLPPPPP | PLMRRRAPLP | PPPPPPLMRR | RAPPPPPPPL | MRRRAPPPPP |
| 70 | 80 | 90 | 100 | 110 | 120 |
| PPPLPRPCSR | PPKTKCSLKP | LHWVKKTRAL | PGSLWDELQR | RQECRDIEDE | QILCAIELSV |
| 130 | 140 | 150 | 160 | 170 | 180 |
| SEIETIFSLG | AKPKPKPEPE | KVPLIDLRRA | TNTEIRLMLL | NIRLPDMIAA | AMAMDESRLD |
| 190 | 200 | 210 | 220 | 230 | 240 |
| DFDQIENLIN | LFPTKEDMKF | LLTYTGDKGN | CEQLFQYLQE | VVKVPRVESK | LRVFSFKIQF |
| 250 | 260 | 270 | 280 | 290 | 300 |
| GTQITKLTKG | LNAVNSACEE | IRTSQKLKDI | MENILCLGNI | LNQGTGRGRA | VGFRLDSLLI |
| 310 | 320 | 330 | 340 | 350 | 360 |
| LSETRADNSK | MTLMHYLCKV | LASKASDLLD | FHKDLESLES | ASKIQLKSLA | EEIQAITKGL |
| 370 | 380 | 390 | 400 | 410 | 420 |
| EKLKQELTAS | ETDGPVSQVF | RKLLKEFISN | AETQVATVMA | LYYPARGNAE | ALAHYFGYHY |
| 430 | 440 | 450 | 460 | ||
| PFEQVTATLL | SFIRLFKKAH | EENVKQAELE | KKKAAKEAEM | EKTK |